
Abstract
Background/Objective
Patients with non-small cell lung cancer (NSCLC) develop resistance to antitumor agents by mechanisms that involve the epithelial-to-mesenchymal transition (EMT). This necessitates the development of new complementary drugs, e.g., cannabinoid receptors (CB1 and CB2) agonists including tetrahydrocannabinol (THC) and cannabidiol (CBD). The combined use of THC and CBD confers greater benefits, as CBD enhances the effects of THC and reduces its psychotropic activity. We assessed the relationship between the expression levels of CB1 and CB2 to the clinical features of a cohort of patients with NSCLC, and the effect of THC and CBD (individually and in combination) on proliferation, EMT and migration in vitro in A549, H460 and H1792 lung cancer cell lines.
Methods
Expression levels of CB1, CB2, EGFR, CDH1, CDH2 and VIM were evaluated by quantitative reverse transcription-polymerase chain reaction. THC and CBD (10–100 μM), individually or in combination (1:1 ratio), were used for in vitro assays. Cell proliferation was determined by BrdU incorporation assay. Morphological changes in the cells were visualized by phase-contrast and fluorescence microscopy. Migration was studied by scratch recolonization induced by 20 ng/ml epidermal growth factor (EGF).
Results
The tumor samples were classified according to the level of expression of CB1, CB2, or both. Patients with high expression levels of CB1, CB2, and CB1/CB2 showed increased survival reaching significance for CB1 and CB1/CB2 (p = 0.035 and 0.025, respectively). Both cannabinoid agonists inhibited the proliferation and expression of EGFR in lung cancer cells, and CBD potentiated the effect of THC. THC and CBD alone or in combination restored the epithelial phenotype, as evidenced by increased expression of CDH1 and reduced expression of CDH2 and VIM, as well as by fluorescence analysis of cellular cytoskeleton. Finally, both cannabinoids reduced the in vitro migration of the three lung cancer cells lines used.





Citation: Milian L, Mata M, Alcacer J, Oliver M, Sancho-Tello M, Martín de Llano JJ, et al. (2020) Cannabinoid receptor expression in non-small cell lung cancer. Effectiveness of tetrahydrocannabinol and cannabidiol inhibiting cell proliferation and epithelial-mesenchymal transition in vitro. PLoS ONE 15(2): e0228909. https://doi.org/10.1371/journal.pone.0228909
Editor: Guillermo Velasco, Universidad Complutense de Madrid, SPAIN
Received: March 28, 2019; Accepted: January 27, 2020; Published: February 12, 2020
Copyright: © 2020 Milian et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability: All relevant data are within the paper.
Funding: This work was supported by grants MAT2016-76039-C4-2-R (MST and CC) and PI16- 01315 (MM) from the Ministry of Economy and Competitiveness of the Spanish Government and the Instituto de Salud Carlos III. CIBER-BBN and CIBERER are funded by the VI National R&D&I Plan 2008-2011, Iniciativa Ingenio 2010, Consolider Program, CIBER Actions and the Instituto de Salud Carlos III, with assistance from the European Regional Development Fund.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Lung cancer is the leading cause of cancer-related death; more than 1 million patients are diagnosed annually. In many cases, life expectancy is only a few months and the 5-year survival rate is < 15% [1–2]. Non-small cell lung cancer (NSCLC) represents 85% of all lung cancers and the most common subtypes are adenocarcinoma and squamous cell carcinoma [3]. Both subtypes are characterized by genetic abnormalities, which lead to alterations in signaling pathways that are targets for drug therapies [4]. All patients with NSCLC eventually develop resistance to antitumor agents, including endothelial growth factor receptor (EGFR) inhibitors and chemotherapeutics, possibly due to abnormal signal transduction and EGFR overexpression [5–7]. This necessitates the development of new complementary pharmacological agents.
The endocannabinoid system is composed of the G-protein–coupled receptors CB1 and CB2, their endogenous ligands anandamide and 2-araquidonoglicerol, and their synthetic and degradative enzymes [8]. CB1 receptor is expressed not only in the central nervous system, but also in other tissues and organs, where its activation exerts both central and peripheral effects [9]. CB2 is expressed in immune cells, microglia, vascular smooth muscle cells, hepatic stellate cells, and endothelial cells. CB2 modulates Ca2+ channels, mitogen-activated protein kinase activation, and cAMP production [9]. According to reports, both receptors are expressed in, for example, breast and prostate cancer, glioblastoma, rhabdomyosarcoma, and colorectal cancer cells [10–14].
Although CB1 and CB2 are expressed in a variety of cancer cell lines and types of tumors, including adenocarcinomas [15], the relationships of their expression levels with lesion characteristics and disease progression have not been investigated. We thus assessed the correlation between the expression levels of the two receptors and the disease and clinical characteristics of a cohort of patients with NSCLC.
Cannabinoid-receptor agonists have potential as complementary pharmacological agents for NSCLC due to their analgesic, antianorexic, antiemetic and antineoplastic properties. For example, cannabinoid receptor agonists modulate key signaling pathways—including the extracellular signal-related kinase (ERK), phosphoinositide 3-kinase (PI3K), p38 mitogen-activated protein kinase (p38 MAPK), and ceramide pathways—in vitro and in vivo, inducing apoptosis and inhibiting cancer dissemination [16–18]. Cannabinoids act on cannabinoid receptors and include endocannabinoids (produced naturally in the body of animals), phytocannabinoids (found in cannabis and some other plants), and synthetic cannabinoids (manufactured artificially). Cannabis sativa contains more than 150 cannabinoid agonists, including Δ9-tetrahydricannabinol (THC), cannabidiol (CBD), cannabinol, cannabichroemene, and cannabigerol [19]. Among these, THC and CBD have demonstrated antitumor efficacy against glioblastoma, leukemia, and melanoma, as well as cervical, breast, and prostate cancer [20]. THC is a partial agonist of CB1 and CB2 receptors, and induces analgesia and muscle relaxation, suppresses emesis and stimulates appetite; however, the psychotropic activity of THC limits its clinical use [21]. CBD has greater affinity for CB2 than CB1 [22]. It also stimulates vanilloid pain receptors (VR1) and inhibits the uptake of anandamide [23]. CBD has anti-inflammatory, neuroprotective, anticonvulsant, muscle-relaxant, and anti-psychotropic effects [22]. Combined used of THC and CBD confers greater benefits, as CBD enhances the effects of THC and reduces its psychotropic activity. Thereby, in rats, CBD administered with THC ameliorate adversely effect (e.g. dysphoria) often associated with THC alone and did not alter the discriminative stimulus effect of THC [24]. Moreover, this combination enhances anticancer activity compared with THC alone and reduces the doses of THC that are needed to inhibit tumor growth [25–27]. CBD has also been shown to alleviate some of the undesired effects of THC administration, such as convulsions, discoordination and psychotic events, and, therefore, improves the tolerance of cannabis-based medicines [25]. Moreover, the combined used of THC and CBD reduces cell viability and migration, and induces apoptosis in human glioblastoma [28]; however, its effect on NSCLC is unclear. We thus investigated the influence of CB1 on the antineoplastic effects of THC in an in vitro model of lung cancer.
The epithelial-to-mesenchymal transition (EMT) involves complex phenotypic changes of tumor cells [29]. During the EMT, epithelial markers (including E-cadherin) are downregulated and mesenchymal markers (such as vimentin [VIM], N-cadherin, and smooth muscle alpha actin [ASMA]) are upregulated by the transcription factors snail, ZEB1, and ZEB2 in a manner involving transforming growth factor-β1 (TGF-β1) [30–32]. Ravi et al. assessed the effect of the CB2 agonist JWH-015 on the EMT in A549 cells exposed to TGF-β1, and in an in vivo model of tumorigenesis [4]. They founded that JWH-015 inhibited EMTE in A549 cells and also reversed the mesenchymal nature of CALU-1 cells by downregulating EGFR signaling. JWH-015 decreased also migratory and invasiveness of A549 cells. In the present study, we also evaluated the effect of the non-selective cannabinoids agonists THC and CBD by separate or in combination on the EMT in three lung cancer cell lines and we explored the additive effect of CBD in combination with THC.
In this study, we investigated the correlation of the expression levels of CB1 and CB2 with the clinical and pathological features of 157 samples of well-characterized lung tumors. We determined the effect of the non-selective cannabinoid agonists CBD and THC by separate or in combination on cell proliferation, the expression of EGFR, the EMT and migration of A549, H460 and H1792 cells exposed to TGF-β or EGF. We found an additive effect of THC/CBD, which support the use of this combination in order to minimize the dose of THC and its psychotropic effects. The results emphasize the importance of the endocannabinoid system, as well as the potential use of CB1 and CB2 as biomarkers of survival in patients with NSCLC.
Materials and methods
Patients
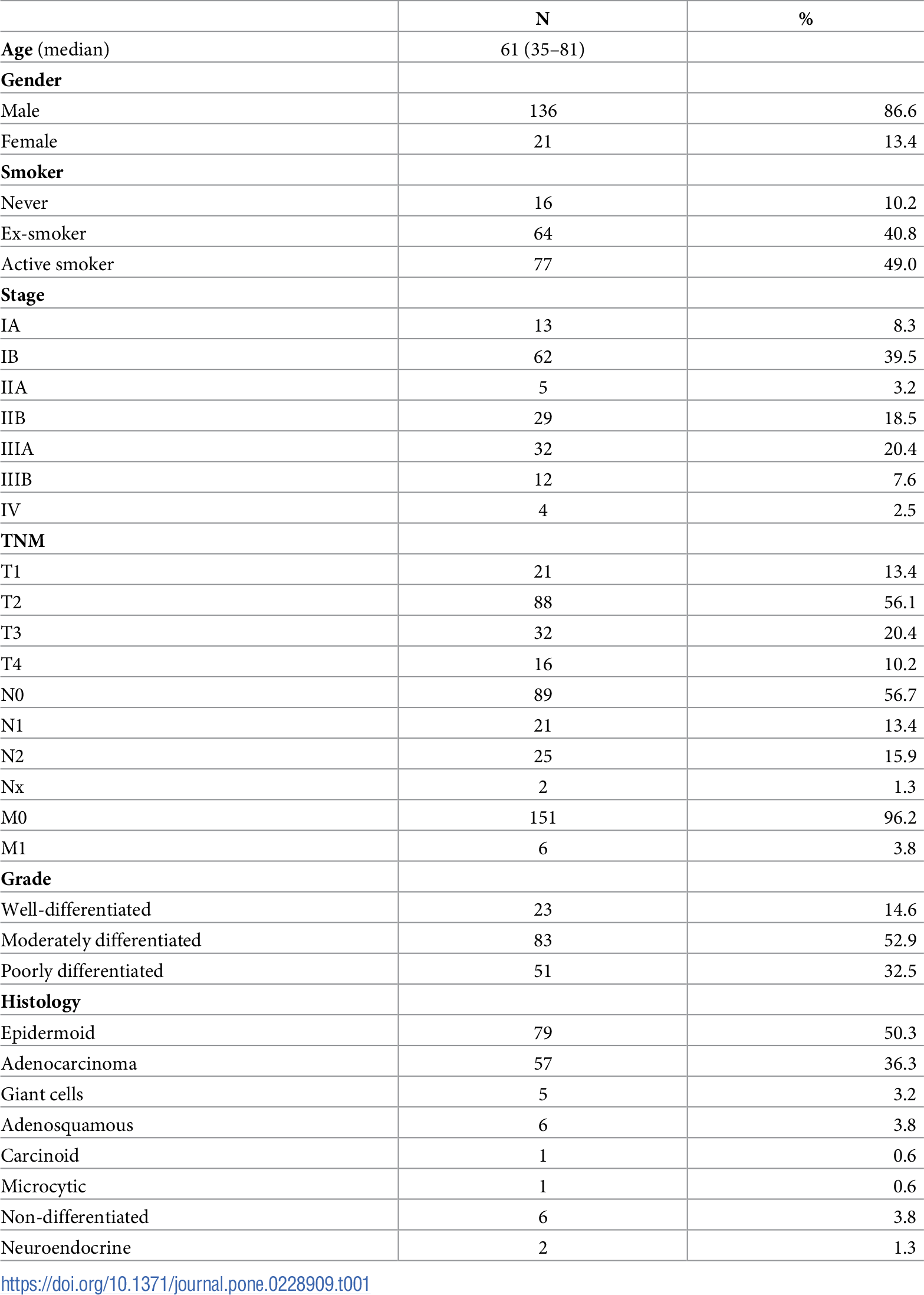
We retrospectively analyzed 157 tumor samples from patients with NSCLC. The tumor samples were stored at −80° C. The clinical and histopathological features of the tumors are listed in Table 1. All of the subjects provided informed consent. The study was conducted in accordance with the Declaration of Helsinki and applicable local regulatory requirements and laws. The procedures were approved by the Ethics Committee of the University General Hospital of Valencia (Spain).

Cannabinoids
THC and CBD were supplied by Sigma-Aldrich (Madrid, Spain). They were prepared at 150 mM amounts in ethanol and used individually or mixed 1:1 at concentrations of 10–100 μM.
Cells and in vitro experiments
A549, H460 and H1792 human lung cancer cells were purchased from the American Type Culture Collection (Rockville, MD, USA). The cells were growth in 25 cm2 culture flasks in Roswell Park Memorial Institute (RPMI) medium containing endotoxin-free fetal bovine serum (FBS, 5% for A549 and H469 cells, 10% for H1792 cells), L-glutamine (4 mM), penicillin (100 U/mL), streptomycin (10 μg/ml) and amphotericin B (0.25 μg/mL).
To assess the antitumor effect of THC and CBD, cells were exposed to 10–100 μM THC or CBD, individually or in combination (1:1 ratio) for 48 hours. Cell proliferation and the expression levels of CB1, CB2, and EGFR were evaluated. To analyze the effect of THC/CBD on the EMT, A549, H460 and H1792 cells were stimulated with TGF-β (15 ng/ml) in the presence or absence of 30 μM THC or 30 μM CBD alone or in combined at 10 μM for 48 hours. The effects on the cytoskeleton and the expression levels of CDH1, CDH2 and VIM were evaluated. Inhibition of the migration of the three cell lines in the presence of 20 ng/ml EGF was also evaluated using the scratch wound assay method.
Cell proliferation assay
The proliferation of A549, H460 and H1792 cells was evaluated by enzyme immunoassay for bromodeoxyuridine (BrdU) incorporation (BrdU Cell Proliferation Assay Kit; Merck Millipore, Darmstadt, Germany). Cells were seeded in 96-well culture plates at 5,000 cells/well and cultured for 24 hours in culture medium as described above. The cells were then cultured for 48 hours in BrdU-containing medium in the presence or absence of 10–100 μM THC and CBD, individually or in combination (1:1 ratio). Culture medium was removed, the cells were fixed, and BrdU incorporation was evaluated with an anti-BrdU antibody by determining the absorbance at 450/550 nm following the manufacturer’s instructions.
Determination of CB1, CB2, EGFR, CDH1, CDH2, and VIM expression levels
Total RNA was extracted from freshly frozen tumor samples and from cell cultures using the TRIzol reagent (Thermo Fischer Scientific Inc., Waltham, MA, USA) according to the manufacturer’s instructions. The RNA concentration was determined by spectrophotometry using a Nanodrop 2000 spectrophotometer (Fischer Scientific, Madrid, Spain). Only extracts with a 260/280 nm ratio > 1.8 were used. RNA integrity was evaluated by capillary electrophoresis using a Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). Only extracts with a RIN of ~10 were used for the determination of gene expression levels.
Random hexamers were used to synthesize complementary DNA (cDNA) using TaqMan RT reagents (Applied Biosystems, Foster City, CA, USA) following the manufacturer’s instructions. Gene expression levels were assayed by reverse transcriptase-polymerase chain reaction (RT-PCR) using Assays on Demand for CB1, CB2, EGFR, CDH1, CDH2, and VIM (Applied Biosystems Madrid, Spain). Reactions were carried out in a 7900HT Real-Time Thermocycler (Applied Biosystems, Madrid, Spain). The comparative ΔΔCt method with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as an endogenous control was used to calculate the relative gene expression levels [33]. For human tissue samples glucuronidase beta (GUSB) gene was used as endogenous control.
Fluorescence staining of F-actin
Filamentous actin (F-actin) in lung cancer cell lines was visualized using rhodamine-conjugated phalloidin (Molecular Probes, Thermo Fisher Scientific, Waltham, MA, USA). Cells were cultured on slides to sub-confluence, and exposed to TGF-β (15 ng/mL) in the presence or absence of 10 μM THC/CBD, which was added 2 hours before TGF-β addition. Control cells received neither TGF-β nor THC/CBD. Forty-eight hours later, the cells were washed twice with pre-warmed phosphate-buffered saline, pH 7.4 (PBS) and fixed in 3.7% formaldehyde in PBS for 10 minutes at room temperature. Next, the cells were permeabilized with 0.1% Triton X-100 in PBS for 3 to 5 minutes. To reduce non-specific background, the samples were pre-incubated with PBS containing 1% bovine serum albumin (BSA) for 20–30 minutes. Next, 5 μL of phalloidin methanolic stock solution were diluted in 200 μL PBS for each sample, and the mixture was added to the samples and incubated for 20 minutes. Finally, the samples were washed, the nuclei were stained with 4′,6-diamidino-2-phenylindole, and the samples were visualized under a DM2500 fluorescence microscope (Leica, Wetzlar, Germany).
Wound healing assay
The wound healing assay was performed as previously described [34]. A549, H460 and H1792 cells were cultured to confluence in six-well culture plates. Then, cultures were scratched to produce a ‘wound’ using sterile 10 μl pipette tips. Cell debris was removed from the culture by washing with sterile PBS for three times. The cells were then cultured in the presence or absence of THC or CBD alone or in combination in serum-free culture medium with 10 ng/ml EGF for 48 hours. Images were recorded using a phase contrast photomicroscope (Nikon), and cell migration was quantified with respect to the control (scratched cultures, non-exposed to EGF cells) using Scion Image software (Alpha 4.0.3.2).
Data analysis
Comparisons of categorical variables were conducted using the non-parametric Mann–Whitney U test or Kruskal–Wallis test. Survival was analyzed by the Kaplan–Meier method and differences between groups were assessed by log-rank test, performed using SPSS software (IBM Corp., Armonk, NY). A p-value of < 0.05 was considered indicative of statistical significance.
In vitro data are shown as means ± SD and were subjected to analysis of variance (ANOVA) followed by Tukey´s multiple-comparison test (GraphPad Software Inc., San Diego, CA, USA). Significance was accepted at p < 0.05. The inhibitory concentration 50 (IC50) of THC and CBD, alone or in combination, was calculated according to proliferation data using the Graphpad software.
Results
CB1 and CB2 overexpression is associated with prolonged survival
First, we evaluated the expression levels of CB1 and CB2 in a well-characterized cohort of patients with NSCLC. The clinical and histopathological features of the patients are summarized in Table 1. The expression levels (ΔCt) of CB1 and CB2 were calculated relative to that of the house-keeping gene glucuronidase beta (GUSB). Multivariate analysis revealed no significant associations between the relative expression levels of CB1 and CB2 and the following clinical characteristics: gender (p = 0.057 and p = 0.267, respectively), progression (p = 0.159, p = 0.209), age (p = 0.690, p = 0.835), smoking (p = 0.223, p = 0.512), stage (p = 0.317, p = 0.961) and histology (p = 0.650, p = 0.550). There was a significant association between chronic obstructive pulmonary disease (COPD) and the expression of CB2 (p = 0.020) but not that of CB1 (p = 0.758).
A survival analysis was performed according to the ΔCt values of CB1 and CB2 (Fig 1). Patients with a ΔCt value of CB1, CB2, or CB1 and CB2 equal to or greater than the mean ΔCt value of the cohort, had longer survival than those with ΔCt values lower than the mean cohort (p = 0.035, 0.126, and 0.025, respectively).

THC/CBD inhibited the proliferation and EGFR expression in lung cancer cells
Next, we assessed the anti-proliferative effects of THC and CBD on A549, H460 and H1792 cells in vitro. The expression of CB1 was significantly higher than that of CB2 (6.58 ± 1.29-fold, n = 6, p = 0.0136; 3.15 ± 1.07-fold, n = 6, p = 0.0160; and 2.15 ± 0.45-fold, n = 6, p = 0.00247, for A549, H460 and H1792 cells, respectively). Because of the cytotoxic effects of both compounds at >100 μM, detected by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, THC and CBD concentrations in the range 10–100 μM were used.
THC and CBD separately or in combination significantly inhibited the proliferation of A549 cells in a dose-dependent fashion (Fig 2A). Concentrations of 10–100 μM of both cannabinoid agonists were studied. IC50 of 27.25 and 37.31 μM were calculated for THC and CBD, respectively. The combination of both cannabinoids significantly reduces the IC50 to 12.94 μM, demonstrating an additive effect. Similar results were founded for H469 cells (IC50 of 30.64, 39.78 and 8.04 μM for THC, CBD or THC/CBD, respectively, Fig 2B) and for H1792 cells (IC50 of 33.39, 46.41 and 14.55 μM for THC, CBD or THC/CBD respectively, Fig 2C). In all cases, the anti-proliferative effect of THC/CBD was inhibited by pertussis toxin (PTX) at 100 ng/mL. According to the calculated IC50 values, a dose of 30 μM for THC and for CBD alone, and 10 μM for THC combined with CBD were used for subsequent experiments.

Both cannabinoid agonists inhibited the EGFR expression in A549, H460 and H1792 cells as it is shown in Fig 2D. Only the combination of THC and CBD significantly decreased EGFR expression in A549 and H460 cells, which was inhibited by PTX at 100 ng/mL.
THC/CBD inhibits the EMT in lung cancer cells
A549, H460 and H1792 cells stimulated with TGF-β were used to assess the effect of THC/CBD on the EMT [4, 31, 32]. Cells were stimulated with 15 ng/ml TGF-β in the presence or absence of 10 μm THC/CBD for 48 hours. Morphological changes were evaluated by phase-contrast light microscopy. Representative results are observed in the Fig 3. Cells exposed to 15 ng /ml TGF-β (Fig 3B, 3J and 3R) became more loosely and acquired a spindle-shaped morphology compared to control cells (Fig 3A, 3I and 3Q). TGF-β-exposed cells treated with 10 μm THC/CBD suppressed these changes (Fig 3C, 3K and 3S). No effect of 10 μM THC/CBD was observed in cells not exposed to TGF-β1 (Fig 3D, 3L and 3T). These effects were found for the three cancer cells included in these experiments. Nevertheless, they were more evident for A549 and H1792 cells than for H460 cells. Changes in cell morphology corresponded to the reorganization of F-actin. In control cells non-exposed to TGF-β or to THC/CBD, phalloidin-labelled F-actin was organized into cortical bundles tightly associated with cell-cell adhesions (Fig 3E, 3M and 3U). After incubation with TGF-β for 48 hours, F-actin was assembled into thicker parallel bundles (actin stress fibers) that crossed the cell surface (Fig 3F, 3N and 3V). These changes were reversed in cultures treated with 10 μM THC/CBD (Fig 3G, 3O and 3W). Interestingly, A549 cells treated with THC/CBD in the absence of TGF-β showed enhanced cell-cell adhesion and thinner actin bundles than control cells (Fig 3H). This effect was not observed for H460 or H1792 cells, in which no per se effect of 10 μM THC/CBD was observed (Fig 3P and 3X).

The changes observed in cell morphology were consistent with those of the expression of the EMT markers CDH1, CDH2 and VIM (Fig 4A–4C). The expression of CDH1 was significantly downregulated (0.25 ± 0.15, 0.30 ± 0.27 and 0.46 ± 0.19-fold for A549, H460 and H1792 cells, respectively), while that of CDH2 (6.45 ± 1.36, 3.86 ± 1.20 and 2.75 ± 0.97-fold for A549, H460 and H1792 cells, respectively) and VIM (7.59 ± 0.69, 4.15 ± 0.20 and 3.91 ± 0.39-fold for A549, H460 and H1792 cells, respectively) were upregulated in cells stimulated with TGF-β compared to control cells. These changes were significantly reversed by CBD or THC alone (30 μM) or in combination (10 μM), which restored the expression values close to those found in treated with CBD or THC but non TGF-β-stimulated cells. The relative expression tendencies found in the three cell types included in this investigation were similar.

Cannabinoids inhibit EGF-induced cell motility in cancer cells
Finally, A549, H460 and H1792 cells were exposed to 20 ng/ml EGF in order to analyze cell migration, as previously reported [34]. As summarized in Fig 5, EGF induces cell migration in the three types of cancer cell used. Both THC and CBD inhibited cell motility, separately or in combination. Although discrete differences were found when comparing data from each cell type, no significant differences were found between them. Inhibition of both cannabinoids tested was close to 30% compared to untreated cells stimulated with EGF.

Discussion
NSCLC is the leading cause of cancer-related mortality in the United States [1–2]. It is an aggressive disease strongly associated with smoking that frequently disseminates; thus, patients are commonly diagnosed with metastatic lesions. Early trials established that radiotherapy was more effective than surgery [35], but due to the extremely low survival rates, chemotherapy has emerged as the optimal modality of treatment. NSCLC is characterized by overactivity of EGFR, which could explain the limited efficacy of chemotherapy [36]. Regrettably, the use of selective EGFR inhibitors or monoclonal antibodies against EGFR has failed, so other therapies for NSCLC are under investigation [36]. Among them, the use of cannabinoids is being evaluated using in vivo and in vitro models of breast, prostate, and lung cancer [37]. Cannabinoids activate the specific G-protein coupled receptors CB1 and CB2. Although CB1 expression has been associated with the brain, and that of CB2 with the immune system, both receptors are expressed to some degree in other cells and organs (e.g., placenta, liver, endothelial cells, smooth muscle cells, and kidney). Although the expression of CB1 and CB2 in NSCLC has been evaluated, to our knowledge, no study has explored the correlation between their expression levels and the clinical and histopathological features of patients with NSCLC. In this study, only COPD was significantly associated with the expression of CB2 (p = 0.020), and non-significantly with that of CB1 (p = 0.758). CB2 is reported to be related to smoking; for example, nicotine addiction is inhibited by CB2 antagonists in mice [38] and the level of CB2 mRNA in blood increases after cessation of marijuana smoking [39]. As smoking is the main causative agent of COPD, further studies of this association are needed to enhance our understanding of the relationship between COPD and lung cancer [40].
The few studies of the prognostic value of CB expression have reported discrepant results [41]. In tumors of the nervous system, there is a relationship between high expression of CB1 and CB2 with astrocytoma, and of CB2 with glioblastoma and malignancy [42–43]. In contrast, high expression of CB2 is associated with an increase of macrophage invasion of brain tumors [44]. In fact, high expression of CB1 is associated with increased severity of prostate and colorectal cancer [45–46]. In contrast, high expression of CB1 and CB2 indicates longer disease-free survival in patients with hepatocellular carcinoma [47], while a lower expression of CB1 is correlated with a lower survival in patients with pancreatic ductal carcinoma [48]. In the present study, NSCLC patients with high expression of CB1 and CB2 showed prolonged survival, which supports their potential use as biomarkers.
Cannabinoids inhibit the proliferation of breast, prostate, and bone cancer cells [31]. However, little is known about the beneficial effects of THC/CBD. Our results indicate that THC and CBD inhibit cell proliferation in a dose-dependent manner. Moreover, CBD enhances the antiproliferative effect of THC in A549, H460 and H1792 cells, in agreement with previous reports [24–27]. CBD and THC also reduce EGFR expression in the three types of cells analyzed in this study. Both cannabinoids are known to inhibit the EGFR pathway, which modulates the proliferation of tumor cells [49–50].
Lung cancer has a propensity to disseminate and invade other tissues [49]. THC inhibits EGFR-induced migration of A549 cells and subcutaneous metastasis in mice with severe combined immunodeficiency [50]. Our results concerning cell migration are in line with those previously reported. Both cannabinoid agonists inhibited EGF-mediated cell migration in A549, H460 and H1792 cells. CBD has an additive effect on the inhibition of THC-mediated cell migration, which supports the beneficial use of both cannabinoids in combination. The metastasis of epithelial tumors involves a series of phenotypic changes, known as the EMT. Cannabinoids have been reported to inhibit EMT in gastric, endometrial and NSCLC cancer cells [4, 34,51]. However, this important property of cannabinoids has not been extensively investigated. For this reason, we explored the effect of THC/CBD on EMT in cancer cells treated with TGF-β in vitro. The combination of THC/CBD strongly affects the cytoskeletal and molecular changes characteristics of EMT, including the downregulation of CDH1 and the upregulation of CDH2 and VIM. Interestingly, in A549 cells, in the absence of TGF-β, exposed to CBD and THC, increased cell-cell adhesion and reduced thickness of actin bundles compared to control cells was observed. These effects could be explained by the mesenchymal phenotype of A549 cells, which are of epithelial origin; indeed, these cells produce TGF-β [52]. Therefore, THC and CBD suppress the basal EMT phenotype, which enhances the medical importance of cannabinoids.
In summary, our results indicate that CB1 and CB2 expression levels have potential as biomarkers for the survival of patients with NSCLC, and that THC and CBD could be used to suppress cell proliferation and EMT. Moreover, the combined use of both compounds could be of interest due to the additive effects observed and could minimize the undesired effects of THC.
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61(2):69–90. pmid:21296855
- 2.Youlden DR, Cramb SM, Baade PD. The international epidemiology of lung cancer: Geographical distribution and secular trends. J Thorac Oncol. 2008;3(8):819–31. pmid:18670299
- 3.Cooper WA, Lam DC, O’Toole SA, Minna JD. Molecular biology of lung cancer. J Thorac Dis. 2013;5 Suppl 5:S479–90.
- 4.Ravi J, Elbaz M, Wani NA, Nasser MW, Ganju RK. Cannabinoid receptor-2 agonist inhibits macrophage induced EMT in non-small cell lung cancer by downregulation of EGFR pathway. Mol Carcinog. 2016;55(12):2063–76. pmid:26741322
- 5.Franklin WA, Veve R, Hirsch FR, Helfrich BA, Bunn PA Jr. Epidermal growth factor receptor family in lung cancer and premalignancy. Semin Oncol. 2002;29(1 Suppl 4):3–14.
- 6.Salomon DS, Brandt R, Ciardiello F, Normanno N. Epidermal growth factor-related peptides and their receptors in human malignancies. Crit Rev Oncol Hematol. 1995;19(3):183–232. pmid:7612182
- 7.Preet A, Qamri Z, Nasser MW, Prasad A, Shilo K, Zou X et al. Cannabinoid receptors, CB1 and CB2, as novel targets for inhibition of non-small cell lung cancer growth and metastasis. Cancer Prev Res (Phila). 2011;4(1):65–75.
- 8.Fowler CJ, Hammarsten P, Bergh A. Tumour cannabinoid CB(1) receptor and phosphorylated epidermal growth factor receptor expression are additive prognostic markers for prostate cancer. PLoS One. 2010;5(12):e15205. pmid:21203460
- 9.Pacher P, Mechoulam R. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Prog Lipid Res. 2011;50(2):193–211. pmid:21295074
- 10.Elbaz M, Nasser MW, Ravi J, Wani NA, Ahirwar DK, Zhao H et al. Modulation of the tumor microenvironment and inhibition of EGF/EGFR pathway: novel anti-tumor mechanisms of cannabidiol in breast cancer. Mol Oncol. 2015;9(4):906–19. pmid:25660577
- 11.Vaccani A, Massi P, Colombo A, Rubino T, Parolaro D. Cannabidiol inhibits human glioma cell migration through a cannabinoid receptor-independent mechanism. Br J Pharmacol. 2005;144(8):1032–6. pmid:15700028
- 12.Jung CK, Kang WK, Park JM, Ahn HJ, Kim SW, Taek Oh S et al. Expression of the cannabinoid type I receptor and prognosis following surgery in colorectal cancer. Oncol Lett. 2013;5(3):870–6. pmid:23426698
- 13.Marshall AD, Lagutina I, Grosveld GC. PAX3-FOXO1 induces cannabinoid receptor 1 to enhance cell invasion and metastasis. Cancer Res. 2011;71(24):7471–80. pmid:22037868
- 14.Cipriano M, Häggström J, Hammarsten P, Fowler CJ. Association between cannabinoid CB₁ receptor expression and Akt signalling in prostate cancer. PLoS One. 2013;8(6):e65798. pmid:23755281
- 15.Ravi J, Sneh A, Shilo K, Nasser MW, Ganju RK. FAAH inhibition enhances anandamide mediated anti-tumorigenic effects in non-small cell lung cancer by downregulating the EGF/EGFR pathway. Oncotarget. 2014;5(9):2475–86. pmid:24811863
- 16.Alexander A, Smith PF, Rosengren RJ. Cannabinoids in the treatment of cancer. Cancer Lett. 2009;285(1):6–12. pmid:19442435
- 17.Bifulco M, Laezza C, Pisanti S, Gazzerro P. Cannabinoids and cancer: Pros and cons of an anti-tumour strategy. Br J Pharmacol. 2006;148(2):123–35. pmid:16501583
- 18.Bifulco M, Laezza C, Portella G, Vitale M, Orlando P, De Petrocellis L et al. Control by the endogenous cannabinoid system of ras oncogene-dependent tumor growth. FASEB J. 2001;15(14):2745–7. pmid:11687506
- 19.Booth JK, Bohlmann J. Terpenes in Cannabis sativa—From plant genome to humans. Plant Sci. 2019; 284:67–72. pmid:31084880
- 20.McAllister SD, Soroceanu L, Desprez PY. The antitumor activity of plant-derived non-psychoactive cannabinoids. J Neuroimmune Pharmacol. 2015;10(2):255–67. pmid:25916739
- 21.Pertwee RG. Neuropharmacology and therapeutic potential of cannabinoids. Addict Biol. 2000;5(1):37–46. pmid:20575818
- 22.Zettl UK, Rommer P, Hipp P, Patejdl R. Evidence for the efficacy and effectiveness of THC-CBD oromucosal spray in symptom management of patients with spasticity due to multiple sclerosis. Ther Adv Neurol Disord. 2016;9(1):9–30.
- 23.Bisogno T, Hanus L, De Petrocellis L, Tchilibon S, Ponde DE, Brandi I et al. Molecular targets for cannabidiol and its synthetic analogues: Effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br J Pharmacol. 2001;134(4):845–52. pmid:11606325
- 24.Vann RE, Gamage TF, Warner JA, Marshall EM, Taylor NL, Martin BR et al. Divergent effects of cannabidiol on the discriminative stimulus and place conditioning effects of delta(9)-tetrahydrocannabinol. Drug Alcohol Depend. 2008;94(1–3):191–8. pmid:18206320
- 25.Velasco G, Sánchez C, Guzmán M. Towards the use of cannabinoids as antitumour agents. Nat Rev Cancer. 2012;12(6):436–44. pmid:22555283
- 26.Massi P, Solinas M, Cinquina V, Parolaro D. Cannabidiol as a potential anticancer drug. Br J Clin Pharmacol. 2013;75(2):303–12. pmid:22506672
- 27.Guindon J, Hohmann AG. The endocannabinoid system and cancer: Therapeutic implications. Br J Pharmacol. 2011;163(7):1447–63. pmid:21410463
- 28.Torres S, Lorente M, Rodríguez-Fornés F, Hernández-Tiedra S, Salazar M, García-Taboada E et al. A combined preclinical therapy of cannabinoids and temozolomide against glioma. Mol Cancer Ther. 2011;10(1):90–103. pmid:21220494
- 29.Kim EJ, Kim HJ, Park MK, Kang GJ, Byun HJ, Lee H et al. Cardamonin suppresses TGF-β1-induced epithelial mesenchymal transition via restoring protein phosphatase 2A expression. Biomol Ther (Seoul). 2015;23(2):141–8.
- 30.Peinado H, Olmeda D, Cano A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat Rev Cancer. 2007;7(6):415–28. pmid:17508028
- 31.Kim JH, Jang YS, Eom KS, Hwang YI, Kang HR, Jang SH et al. Transforming growth factor beta1 induces epithelial-to-mesenchymal transition of A549 cells. J Korean Med Sci. 2007;22(5):898–904. pmid:17982242
- 32.Kawata M, Koinuma D, Ogami T, Umezawa K, Iwata C, Watabe Tet al. TGF-β-induced epithelial-mesenchymal transition of A549 lung adenocarcinoma cells is enhanced by pro-inflammatory cytokines derived from RAW 264.7 macrophage cells. J Biochem. 2012;151(2):205–16. pmid:22161143
- 33.Mata M, Sarriá B, Buenestado A, Cortijo J, Cerda M, Morcillo EJ. Phosphodiesterase 4 inhibition decreases MUC5AC expression induced by epidermal growth factor in human epithelial cells. Thorax. 2005;60(2):144–52. pmid:15681504
- 34.Xian X, Huang L, Zhang B, Wu C, Cui J, Wang Z. WIN 55,212–2 inhibits the epithelial mesenchymal transition of gastric cancer cells via COX-2 signals. Cell Physiol Biochem. 2016;39(6):2149–57. pmid:27802436
- 35.Zappa C, Mousa SA. Non-small cell lung cancer: Current treatment and future advances. Transl Lung Cancer Res. 2016;5(3):288–300. pmid:27413711
- 36.Bethune G, Bethune D, Ridgway N, Xu Z. Epidermal growth factor receptor (EGFR) in lung cancer: An overview and update. J Thorac Dis. 2010;2(1):48–51. pmid:22263017
- 37.Hermanson DJ, Marnett LJ. Cannabinoids, endocannabinoids, and cancer. Cancer Metastasis Rev. 2011;30(3–4):599–612. pmid:22038019
- 38.Ignatowska-Jankowska BM, Muldoon PP, Lichtman AH, Damaj MI. The cannabinoid CB2 receptor is necessary for nicotine-conditioned place preference, but not other behavioral effects of nicotine in mice. Psychopharmacology (Berl). 2013;229(4):591–601.
- 39.Muhl D, Kathmann M, Hoyer C, Kranaster L, Hellmich M, Gerth CW et al. Increased CB2 mRNA and anandamide in human blood after cessation of cannabis abuse. Naunyn Schmiedebergs Arch Pharmacol. 2014;387(7):691–5. pmid:24788457
- 40.Mouronte-Roibás C, Leiro-Fernández V, Fernández-Villar A, Botana-Rial M, Ramos-Hernández C, Ruano-Ravina A. COPD, emphysema and the onset of lung cancer. A systematic review. Cancer Lett. 2016;382(2):240–4. pmid:27666776
- 41.Pyszniak M, Tabarkiewicz J, Łuszczki JJ. Endocannabinoid system as a regulator of tumor cell malignancy—biological pathways and clinical significance. Onco Targets Ther. 2016;9:4323–36. pmid:27486335
- 42.Sánchez C, de Ceballos ML, Gomez del Pulgar T, Rueda D, Corbacho C, Velasco G et al. Inhibition of glioma growth in vivo by selective activation of the CB(2) cannabinoid receptor. Cancer Res. 2001;61(15):5784–9. pmid:11479216
- 43.Ellert-Miklaszewska A, Grajkowska W, Gabrusiewicz K, Kaminska B, Konarska L. Distinctive pattern of cannabinoid receptor type II (CB2) expression in adult and pediatric brain tumors. Brain Res. 2007;1137(1):161–9. pmid:17239827
- 44.De Jesús ML, Hostalot C, Garibi JM, Sallés J, Meana JJ, Callado LF. Opposite changes in cannabinoid CB1 and CB2 receptor expression in human gliomas. Neurochem Int. 2010;56(6–7):829–33. pmid:20307616
- 45.Gustafsson SB, Palmqvist R, Henriksson ML, Dahlin AM, Edin S, Jacobsson SO et al. High tumour cannabinoid CB1 receptor immunoreactivity negatively impacts disease-specific survival in stage II microsatellite stable colorectal cancer. PLoS One. 2011;6(8):e23003. pmid:21901119
- 46.Chung SC, Hammarsten P, Josefsson A, Stattin P, Granfors T, Egevad L et al. A high cannabinoid CB(1) receptor immunoreactivity is associated with disease severity and outcome in prostate cancer. Eur J Cancer. 2009;45(1):174–82. pmid:19056257
- 47.Xu X, Liu Y, Huang S, Liu G, Xie C, Zhou J et al. Overexpression of cannabinoid receptors CB1 and CB2 correlates with improved prognosis of patients with hepatocellular carcinoma. Cancer Genet Cytogenet. 2006;171(1):31–8. pmid:17074588
- 48.Michalski CW, Oti FE, Erkan M, Sauliunaite D, Bergmann F, Pacher P et al. Cannabinoids in pancreatic cancer: Correlation with survival and pain. Int J Cancer. 2008;122(4):742–50. pmid:17943729
- 49.Elbaz M, Nasser MW, Ravi J, Wani NA, Ahirwar DK, Zhao H et al. Modulation of the tumor microenvironment and inhibition of EGF/EGFR pathway: Novel anti-tumor mechanisms of cannabidiol in breast cancer. Mol Oncol. 2015;9(4):906–19. pmid:25660577
- 50.Preet A, Ganju RK, Groopman JE. Delta9-tetrahydrocannabinol inhibits epithelial growth factor-induced lung cancer cell migration in vitro as well as its growth and metastasis in vivo. Oncogene. 2008;27(3):339–46. pmid:17621270
- 51.Zhang Y, Zheng W, Shen K, Shen W. Δ9-Tetrahydrocannabinol inhibits epithelial-mesenchymal transition and metastasis by targeting matrix metalloproteinase-9 in endometrial cancer. Oncol Lett. 2018;15(6):8527–35. pmid:29805589
- 52.Koyama S, Sato E, Nomura H, Kubo K, Nagai S, Izumi T. Type II pneumocytes release chemoattractant activity for monocytes constitutively. Am J Physiol. 1997;272(5 Pt 1):L830–7.

